Between November 5th and November
8th, I listened to over 120 talks about the latest research in paleontology. I
was at the Society of Vertebrate Paleontology's 74th Annual Meeting in Berlin,
Germany. Below is a highlights reel of what I learned at this year’s
conference.
Evolution
of Flight
 A major theme of this year’s talks was figuring
out how the modern suite of features seen in birds came into existence. We know
birds branched from the theropod dinosaur lineage, thanks to feathered fossils
such as Archaeopteryx, but the
details are still being sorted out as to how each step of the transition
occurred. The most famous specimen of Archaeopteryx
is housed at Berlin’s Museum für Naturkunde, hence the focus of this year’s
conference on this issue.
A major theme of this year’s talks was figuring
out how the modern suite of features seen in birds came into existence. We know
birds branched from the theropod dinosaur lineage, thanks to feathered fossils
such as Archaeopteryx, but the
details are still being sorted out as to how each step of the transition
occurred. The most famous specimen of Archaeopteryx
is housed at Berlin’s Museum für Naturkunde, hence the focus of this year’s
conference on this issue.
Previously held
beliefs are being overturned. For example, a mouthful of teeth was not a
hindrance to flying—plenty of dinosaurs in the fossil record had toothed jaws
(rather than beaks) and had a
skeleton fully capable of flight. Likewise, not
having teeth (having a beak instead), does not seem to indicate any
improvement in the skeleton for flight. So while changing from a toothed mouth
to a beak would certainly lighten up a skull for flying, the initial teeth-to-beak
change may have been diet-related, and certainly did not predate flight.
Modern birds have shortened tails, while
early birds did not. For a long time, paleontologists assumed long tails meant
an animal was not a great flyer. However, new studies have shown that long
tails can help with flight stabilization, and that wide long tails are quite useful in lift. Long tails remained “in
vogue” for millions of years during the evolution of flight, and were possibly
related to the launching mechanism of early birds.
The idea above has a lot of punch
because the back legs of ancient birds were not necessarily well developed for
launching birds into the air. In fact, in many cases, the hind limbs were
actually used in flying—as wings. There
is a case to be made that having four wings is actually ancestral to having
two. That is, birds evolved from four-winged ancestors, and their back legs
eventually lost their flight capabilities in favor of other uses, such as
helping with bipedal gripping in trees, hunting/fighting, and eventually,
launching birds from the ground up.
Along this line of thought, a great deal
of research suggests Archaeopteryx
was transitional, not just between non-avian dinosaurs and birds, but between
four-winged fliers and two-winged fliers. There are feathers on its hind limbs
that resemble flight feathers, but would not be strong enough for serious use.
On its forelimbs, the feathers are not stable enough for sustained flight by
themselves, though they are better than what is found on the hind limbs. One
speaker suggested a “parkour” model for how Archaeopteryx
got around. It could launch itself and move short distances in the air, but
it was not going to fly for long periods of time. It had perhaps secondarily
lost its ability to fly. However, this loss of flying ability freed up its back
legs, which eventually helped its descendants
become better flyers.
A final note of controversy to all of
this: a new study showed that Archaeopteryx
was much slower growing than modern birds, and took over a full year to
mature. All the specimens we have are from young individuals, and therefore
there is a chance that, just like a fledgling bird today, perhaps the Archaeopteryx skeletons on record only
appear to belong to poor flyer because they had yet to fully mature.
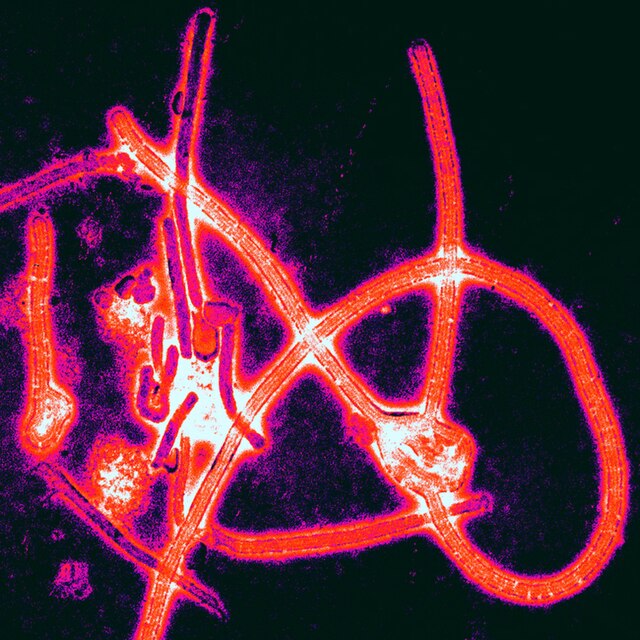
Colors in Dinosaurs
 |
Possible
fossil melanosomes. Credit: Holly Barden.
|
Finding evidence
of color in feathers is one of the most exciting things to have happened in
paleontology in recent years. However, fossil melanosomes (which are what tell
us about color of feathers) can easily be confused with fossil bacteria (which would
have moved in when the animal died, and have nothing to do with the color). The
shapes are very similar, so appearance alone would not be enough to distinguish
them. But what else is there to go on when all we have are the fossil
impressions?
One talk at the conference proposed a
solution to this problem: look at the matrix that the fossils are embedded in.
Beta keratin, another structure found in feathers, can survive extreme
degradation, and actually outlast melanosomes in the decaying process.
Meanwhile, bacteria survive on biofilm, which is visually distinct from beta
keratin. If we can detect beta keratin in the sediment matrix, we can be
reasonably assured we are looking at melanosomes. If we just have a mess of
biofilm, and no beta keratin, there is zero chance melanosomes have survived,
and therefore the structures must be bacterial.
Spinosaurus
 |
Drawings
of the destroyed bones. 1915.
|
This giant theropod dinosaur is famous
for its sailed back and tragic history (also its questionable role in Jurassic
Park III). The lower jaw and tall spines of its back were discovered in the early
20th century and sent to Munich to be stored in the Barvarian State
Collection Museum. This museum was demolished in WWII by allied bombing, and Spinosaurus was lost to science.
In 2008, a century after its initial discovery, some
fossil fragments were found—including the trademark spines. These fragments
were shown to Dr. Ibrahim by a random man in a desert town in Morocco. Years
later, an Italian fossil collector contacted Dr. Ibrahim with a whole slew of
bones that were quickly shown to also be Spinosaurus.
They were similar in their matrix sediments to the fragments Dr. Ibrahim
had been shown before, and had been bought through some sketchy black market
trades from an “unknown” collector. Dr. Ibrahim put two and two together.
Clearly, the mystery man from Morocco knew where to find Spinosaurus, but no one knew where to find him. Dr. Ibrahim spent months going from tiny town to tiny town
trying to track this man down, but the only clue he had was that this man had a
mustache—which didn’t really narrow down the search in Morocco.
At lunch with colleagues, planning the
details for his flight back home and regretting that he would have to give up
the search, Dr. Ibrahim saw a familiar looking face walk by. It was the mystery
man! Long story short, the man agreed to show them to the site where he’d been
digging up bones, and Ibrahim’s team found the rest of what remained of Spinosaurus.
This discovery showed that not only is Spinosaurus the longest known predatory
dinosaur (at 15 meters), it is also possibly semi-aquatic. Its limb proportions
are similar to that found in modern-day semi-aquatic animals, and its wide flat
feet indicate it could have used them for swimming. Also, its bones have
condensed cross sections, much like penguins have. Its teeth and jaw were
shaped for catching and eating fish, and it probably ate really big fish. This is the first evidence of
a non-avian dinosaur having adaptations for the water!
Did
Giant Kangaroos Hop or Walk?
In what might be the silliest sounding
debate in paleontology, we have this question.
Giant Pleistocene Kangaroos reached sizes over 500lbs, and the debate is as to
whether or not they could have hopped at this extremely large size without
breaking their legs. The stress on the bones is super high in these larger
animals, but by looking at modern kangaroos, it turns out that the time of the
stride matters more than the mass of the animal in terms of hopping abilities. So
if this kangaroo moved fast enough, it could, conceivably, hop. It has the
proper limb ratio to allow for hopping, after all. However, its robust hips,
knees, and ankles suggest it could have walked around as other animals do,
supporting its weight on one limb at a time. Did it walk all the time? Did it
hop only at fast speeds? How would it build up to that? This is a fantastic,
ongoing debate that I cannot wait to see discussed further.
Dinosaur
Parenting
Oviraptors, once thought to be egg-stealing
predators, are actually great parents. They have been found near eggs because
they guard their eggs. This has been
known for quite some time, but what isn’t known is what their brooding behavior
was like, or if they were even true brooders.
In modern birds and crocodilians,
eggshells are porous in nests that are covered, but have low porosity in nests
that are open-air and brooded by parents. All oviraptor species tested have
low-porosity eggs, which indicates their eggs were kept in open-air nests and
that the dinosaurs likely brooded. The small-to-medium sized species had eggs
that could withstand their parents’ body mass (at least when that mass is spread
over the full clutch of eggs), but some oviraptor species were large enough to
squash their eggs. So how could they possibly brood?
 |
| Bad drawings by me. |
The answer lies
in their nest design. A study comparing the inner diameter to the outer
diameter of nests showed that small-to-medium sized oviraptors packed their
eggs in close, while larger oviraptors had a large central open space in their
nest, so the parent could hang out in the middle without squashing anyone. This
shows that oviraptors had behavioral differences to ensure they could brood,
indicating how important this practice was to them.
And for the first time, there is nearly
indisputable evidence that some dinosaurs used the same nesting site from year
to year. Troodontids—related to oviraptors—had clutches of eggs nearly on top
of one another. Each clutch was successfully hatched, and the incredibly close
proximity suggests that it was the same parent using the site again and again.
Other dinosaurs have been shown to reuse nesting grounds, but until now it was
never clear if it is the same individuals reusing the site, or others of that species
reusing the site. This dedication to a single nest might also indicate that
mated pairs stayed together for longer than one season, since this is the kind
of behavior seen in many birds who “mate for life”.
Environmental
Changes and Animal Response
The fossil record is incredibly useful
for understanding the link between animals and their environment. This is of
particular importance in today’s modern world, as humans continue to change
habitats globally.
Amphibians are great indicator species
for climate change. My favorite talk that demonstrated this was a study on fossil
tiger salamanders. They were shown to be less neotonic during glacial periods,
and more likely to exist in their metamorph forms during interglacial periods,
when the environment was less predictable. This can now be used in looking out
how tiger salamanders respond to changes in the world today.
A study also looked at the timing and
circumstances of the extinction of the giant shark “Megalodon” (Carcharocles megalodon) to better
understand what is happening to the large shark die-off happening today.
Megalodon did not appear to be increasing in size leading to its extinction, so
growing “too big for its britches” was not what did it in (as has been
suggested before). The extinction seems to have taken place around 2.5 million
years ago, and coincides with the extinction of much of its smaller bodied prey
and migration of other kinds of ocean predators. This demonstrates an ancient
food chain collapse, and predicts what will happen in our modern oceans today.
On the flip side, not all extinctions
can be linked to rapid environmental change. European apes went extinct around
9 million years ago, and for a long time their extinction has been connected to
a “cooling crisis” that rapidly shifted the environment from forests to
grasslands. However, a new study found that grasslands did not increase in
these areas during this time, and temperatures did not drop nearly as
drastically as previous studies had shown. So this extinction remains a mystery.
In fact, in some cases, animal
extinctions may trigger changes in the environment, rather than the other way
around. This was the case when mastodons died off at the end of the Pleistocene.
At this time, hardwood trees made a huge comeback and forests increased, since
their biggest “predator” was extinct.
Finally, it may be time to rethink how
and why animals diversify. The leading mentality behind diversification has
always been that a new innovation will drive the origination of new species,
such as the origin of feathered flight causing the explosion of bird species
that we see today. However, looking closer, it becomes clear that while
innovations help species survive extinction events, it is only after the extinction event that true
diversification occurs. To state this even more simply: extinction drives
speciation and innovation merely sets the stage.
Along these lines, the “Out of Tibet”
hypothesis is gaining traction. Before the big Pleistocene Ice Age, the Tibetan
Plateau was a “training ground” for cold-adapted features on animals. Due to
its high elevation, living on the Tibetan Plateau is like living in the arctic,
and once the Ice Age hit and the environment changed, animals already
cold-adapted had a field day (while others around the world died off). For
example, it’s well known that the Woolly Rhino definitely migrated out of Tibet
during the Ice Age. Now there is new evidence that the ancestors of artic
foxes, bighorn sheep, and even bison may have done so as well. Meanwhile, there
is also new fossil evidence of snow leopards and blue sheep on the Tibetan
Plateau. Their descendants still exist there today, and this traces their
predator/prey relationship back millions of years—an extremely rare connection
to be able to make.
Other
Exciting Finds
- A
new protocetid (ancient whale) from Egypt that may show some of the transition
between foot powered swimming to tail powered swimming.
- A
new theropod dinosaur, Tachiraptor,
is Venezuela’s second-ever discovered dinosaur.
-
Fossil
adult tiger salamanders were found in a mammoth tusk—they were possibly using
it as a shelter!
- A
pregnant mare of an ancient horse species (Eurohippus
messelenses) was found in the Messel Pit in Germany, with soft tissue
impressions left on the fossil, including the broad ligament, uterus, and
placenta.
- A
new dinosaur was found in Antarctica! It looks related to the Iguanodon clade, but is still being
described and has no name yet. Only four named species of dinosaur have ever
been found in Antarctica.
- A
new early ceratopsian (relative of Triceratops)
from China will shed light on that whole group’s evolution.
- A
new ichthyosaur from the Jurassic of Germany suggests that shorter, broader
flippers may have been the evolutionary direction this lineage was heading in.
- And
finally, there is something super cool that I have
been sworn to secrecy on—sorry! Hopefully I'll get to blog about this one later.
Other
New Studies
- A
look at the inner ear of snakes has scored points for the hypothesis that they
came from terrestrial ancestors who likely tunneled in the earth, rather than
from ancestors evolving to fit an aquatic habitat. Both have been leading
hypotheses in why snakes lost their legs.
-
Looking
at the inner ear of sauropod dinosaurs (long-necked dinosaurs) shows an
evolutionary trend towards sweeping neck movements rather than raising their
necks up and down. This has implications on their feeding behaviors.
- Pinning
down the origin of sea turtles has always been difficult, but a new study
suggests they came from a single lineage in the Late Jurassic, and later
developed into giant-sized species multiple times, independently (through
convergent evolution).
- Around
35 million years ago, many species managed to cross the Tethys Sea to colonize
Africa from Asia. The most common of these were anthropoid primates and
hystricognathian rodents. In fact, some of these species then also went on to
colonize South America from Africa. This shows that these cross-sea
colonizations were non-random, and that certain species managed them
repeatedly. What these species have in common are small body sizes, climbing
skills, and group life, which supports the rafting hypothesis of colonization.
(Animals clung to clumps of vegetation that moved across the sea after being
swept out in a major storm.)
- Growth
patterns on Titanoboa bones show that
it grew faster than other snakes, and for a longer time than other snakes. A
double whammy to explain its huge size.
-
Ankylosaurs
had a bony hyobranchium (part of the neck region), rather than a cartilaginous
one. This makes for stronger muscle attachments for the tongue, and indicates
some sort of specialized food processing technique not seen in other animals. The
question remains: what?
-
Tiktaalik had reduced
skull movements in feeding, intermediate between the stiff skulls of tetrapods
and the bendy/mobile skulls of fish. Which makes beautiful sense, given its
placement as a transitionary animal!
- The
switch to breathing with lungs rather than gills may have less to do with
transitioning to non-marine habitats, and more to do with surviving in marine
habitats that occasionally experienced hypoxic conditions.
- Plesiosaur
taxonomy (relationships) may need to be re-thought, because no one has taken
into account that plesiosaurs gave live birth and therefore had dimorphic
pelvic bones. What some people are calling different species might just be
different sexes of the same species!
There were also tons of talks on
methodology, new fossil localities, mammalian evolution (my personal favorite), and much more. I have
copious notes beyond what is presented here, so feel free to contact me if you have
further questions. To anyone whose research I presented: I tried to only present talks that did not request social-media-silence. If you see something here you would like corrected or removed, please contact me! If I did not present your research and you would like me to in the future, also feel free to contact me. The summary above is not meant to slight anyone's work, and is only meant to translate a few tidbits for non-academic audiences. Also, keep in mind that certain topics and talks I am saving for future posts where I can delve into them in more detail.
CITATIONS
Barnosky,
A., Lindsey, E., Villavicencio, N., Marshall, C. FOSSIL EVIDENCE FOR LASTING
ECOLOGICAL TRANSFORMATION AS A RESULT OF DEFAUNATION. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 85.
Barrett,
P., Milner, A., Hooker, J. A NEW ORNITHOPOD DINOSAUR FROM THE LATEST CRETACEOUS
OF THE ANTARCTIC PENINSULA. Journal of Vertebrate Paleontology, Program
and Abstracts, 2014, Page 85.
Beard,
K., Coster, P., Marivaux, L., Salem, M., Chaimanee, Y., Jaeger, J. NONRANDOM
BUT UNPARSIMONIOUS PATTERNS OF MAMMALIAN DISPERSAL BETWEEN ASIA AND AFRICA
DURING THE LATER PALEOGENE. Journal of Vertebrate Paleontology, Program
and Abstracts, 2014, Page 87.
Brocklehurst,
N. INNOVATION, EXTINCTION AND RATES OF CLADOGENESIS IN EARLY AMNIOTES. Journal
of Vertebrate Paleontology, Program and Abstracts, 2014, Page 98.
Casanovas,
V., Kimura, Y., Flynn, L., Alba, D., Pilbeam, D., Moya-Sola, S. THE END OF THE
MIOCENE HOMINOID RADIATION IN EURASIA: NEW INSIGHTS PROVIDED BY STABLE ISOTOPE
ANALYSIS OF TOOTH ENAMEL IN MUROID RODENTS. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 105.
Clement,
A., Ahlberg, P. BUCCAL-PUMP AIR GULPING IN DEVONIAN LUNGFISHES (SARCOPTERYGII;
DIPNOI): INSIGHTS FROM TOMOGRAPHIC DATA. Journal of Vertebrate Paleontology,
Program and Abstracts, 2014, Page 110.
Cook,
J., Attard, M., Krall, P., Wroe, S. COULD THE GIANT KANGAROO HOP? Journal of
Vertebrate Paleontology, Program and Abstracts, 2014, Page 112.
Dyke, G., Cau, A., Naish, D.,
Brougham, T., Godefroit, P. ARCHAEOPTERYX
AND PARAVIAN PHYLOGENY: THE ENIGMA OF BALAUR.
Journal
of Vertebrate Paleontology, Program and Abstracts, 2014, Page 123.
Erickson,
G., Rauhut, O., Roeper, M., Lochner, E., Norell, M. LONG BONE HISTOLOGY,
GROWTH, AND THE 'BLOOD' OF ARCHAEOPTERYX.
Journal of Vertebrate Paleontology, Program and Abstracts, 2014, Page
126.
Franzen,
J., Aurich, C., Habersetzer, J. A PREGNANT MARE WITH FETUS OF EUROHIPPUS MESSELENSIS (MAMMALIA,
PERISSODACTLYA, EQUIDAE) FROM THE EARLY MIDDLE EOCENE OF MESSEL PIT (GERMANY). Journal
of Vertebrate Paleontology, Program and Abstracts, 2014, Page 132.
Gingerich,
P., Antar, M., and Zalmout, I. SKELETON OF A NEW PROTOCETID (CETACEA,
ARCHAEOCETI) FROM THE LOWER GEHANNAM FORMATION OF WADI AL HITAN IN EGYPT: SURVIVAL
OF A PROTOCETID INTO THE PRIABONIAN LATE EOCENE. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 138.
Habib,
M. RECONSTRUCTING LOCOMOTOR PERFORMANCE IN ARCHAEOPTERYX.
Journal of Vertebrate Paleontology, Program and Abstracts, 2014, Page 143.
Hill,
R., D’Emic, M., Bever, G., Norell, M. THE MOST COMPLETE, OSSIFIED HYOBRANCHIAL
APPARATUS OF A FOSSIL DINOSAUR: IMPLICATIONS FOR ONTOGENY AND FUNCTIONAL ANATOMY
Journal of Vertebrate Paleontology, Program and Abstracts, 2014, Page 147.
Ibrahim,
N., Maganuco, S., Sereno, P., Dal Sasso, C., Keillor, T., Martill, D., Zouhri,
S., Fabbri, M., Auditore, M. ASSOCIATED REMAINS OF SPINOSAURUS AEGYPTIACUS, AN ENORMOUS PREDATORY DINOSAUR WITH
SUBAQUATIC ADAPTATIONS. Journal of Vertebrate Paleontology, Program and
Abstracts, 2014, Page 151.
Janis,
C., Kuchenbecker, K., Figueirido, B. LOCOMOTION IN EXTINCT GIANT KANGAROOS:
WERE STHENURINES HOP-LESS MONSTERS? Journal of Vertebrate Paleontology,
Program and Abstracts, 2014, Page 153.
Lemberg,
J., Ross, C., Shubin, N., Daeschler, E. FUNCTIONAL IMPLICATIONS OF PLATYROSTRAL
SKULLS FOR FEEDING IN WATER WITH INSIGHTS INTO THE CRANIAL MORPHOLOGY OF TIKTAALIK ROSEAE. Journal of
Vertebrate Paleontology, Program and Abstracts, 2014, Page 168.
Longrich,
N. PRIMITIVE FEATHER ARRANGEMENT IN ARCHAEOPTERYX
LITHOGRAPHICA SHEDS LIGHT ON THE ORIGIN AND EVOLUTION OF BIRDS. Journal
of Vertebrate Paleontology, Program and Abstracts, 2014, Page 170.
Maxwell,
E., Fernandez, M., Larsson, H., Schoch, R. A NEW ICHTHYOSAUR FROM THE MIDDLE
JURASSIC OF GERMANY PROVIDES INSIGHTS INTO THE ASSOCIATION BETWEEN LIMB FORM AND
FUNCTION. Journal of Vertebrate Paleontology, Program and Abstracts,
2014, Page 183.
Moyer,
A., Zheng, W., Schweitzer, M. USING DEGRADATION EXPERIMENTS OF EXTANT FEATHERS
TO ADDRESS THE PRESENCE OF MICROBODIES IN FOSSIL FEATHERS. Journal of
Vertebrate Paleontology, Program and Abstracts, 2014, Page 192.
O’Conner, J. and Zhou, Z. EARLIEST
STAGES IN THE EVOLUTION OF THE MODERN AVIAN SKELETON: ARCHAEOPTERYX AND THE JEHOL AVIFAUNA COMPARED. Journal of Vertebrate
Paleontology,
Program and Abstracts, 2014, Page 197.
Pimiento,
C. EXTINCTION AND BODY SIZE PATTERNS OF THE GIANT SHARK CARCHAROCLES MEGALODON. Journal of Vertebrate Paleontology,
Program and Abstracts, 2014, Page 205.
Rabi,
M. ORIGIN AND EARLY EVOLUTION OF SEA TURTLES. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 210.
Rauhut,
O. and Foth, C. NEW INFORMATION ON THE THEROPOD DINOSAURS FROM THE LATE JURASSIC
LITHOGRAPHIC LIMESTONES OF SOUTHERN GERMANY. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 212.
Schmitt,
A. ACCELERATING OUR UNDERSTANDING OF THE INNER EAR OF SAUROPODOMORPHA: FIRST GLOBAL,
STATISTICAL ANALYSIS OF SEMICIRCULAR CANALS OF DIPLODOCID AND MACRONARIAN DINOSAURS
AND ITS IMPLICATION FOR NECK-POSTURE. Journal of Vertebrate Paleontology,
Program and Abstracts, 2014, Page 223.
Schumacher,
B. PHYLOGENY AND POSITED DIMORPHISM OF POLYCOTYLID PLESIOSAURS, NEW INFORMATION
FROM A NEARLY COMPLETE SKELETON OF POLYCOTYLUS
LATIPINNIS FROM THE NIOBRARA FORMATION (EARLY CAMPANIAN) OF SOUTH DAKOTA,
UNITED STATES. Journal of Vertebrate Paleontology, Program and
Abstracts, 2014, Page 225.
Sellers,
W. and Manning, P. THE APPLICATIONS OF EVOLUTIONARY ROBOTICS TO RECONSTRUCTING
LOCOMOTION IN EXTINCT ANIMALS. Journal of Vertebrate Paleontology,
Program and Abstracts, 2014, Page 227.
Tanaka,
K., Zelenitsky, D., Lü, J., Yi, L, Pu, H., Chang, H., Xu, L.,
Li, H. NEST
TYPE AND INCUBATION BEHAVIOR IN OVIRAPTOROSAURS IN RELATION TO BODY SIZE. Journal
of Vertebrate Paleontology, Program and Abstracts, 2014, Page 238.
Varricchio,
D., Jackson, F., Jin, X. LAY-BROOD-REPEAT: NESTING SITE FIDELITY IN ECOLOGIC
TIME FOR TWO CRETACEOUS TROODONTID DINOSAURS. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 246.
Wang,
X., Li, Q., Tseng, Z., Takeuchi, G., Deng, T. THE PLIOCENE TIBETAN PLATEAU AS A
TRAINING GROUND FOR COLD ENVIRONMENT ADAPTATION AND ORIGIN OF HOLARCTIC MEGAFAUNA.
Journal of Vertebrate Paleontology, Program and Abstracts, 2014, Page
251.
Werning,
S., Head, J., Bloch, J. BONE HISTOLOGY AND GROWTH IN THE LARGEST KNOWN SNAKE, TITANOBOA CERREJONENSIS. Journal of
Vertebrate Paleontology, Program and Abstracts, 2014, Page 254.
Yi,
H. RESOLVING THE LOCOMOTORY ECOLOGY OF THE ANCESTRAL SNAKE: LISTENING TO WHAT
THE EAR TELLS US. Journal of Vertebrate Paleontology, Program and
Abstracts, 2014, Page 259.
Zhang,
Z. and Hadly, E. LIFE
HISTORY DYNAMICS OF THE TIGER SALAMANDER, AMBYSTOMA
TIGRINUM, IN RESPONSE TO LATE PLEISTOCENE CLIMATE. Journal of Vertebrate
Paleontology, Program and Abstracts, 2014, Page 260.
Zheng,
W., Jin, X., Xu, X. A NEW BASAL NEOCERATOPSIAN (ORNITHISCHIA, CERATOPSIA) FROM THE
LATE CRETACEOUS OF CENTRAL CHINA. Journal of Vertebrate Paleontology,
Program and Abstracts, 2014, Page 260.


.jpg/1024px-Signal_received_from_Rosetta_(12055070794).jpg)



{kind=link}